【連載】エクソソームと生命現象「第 3 回 細胞老化とエクソソーム」
本記事は、和光純薬時報 Vol.86 No.4(2018年10月号)において、公益財団法人がん研究会 がん研究所 細胞老化プロジェクト 岡田 遼様、国立研究開発法人科学技術振興機構 さきがけ 高橋 暁子様に執筆いただいたものです。


はじめに
日本人の平均寿命は年々延長し続けており、現在では我が国は世界有数の長寿国として知られている。その一方で、がん・動脈硬化・アルツハイマー・肺線維症・骨粗鬆症のように加齢と共に罹患率が上昇する疾患(加齢性疾患)の増加が深刻な社会問題となっている。
加齢性疾患の発症には様々な複合的な要因があげられるが、その原因の一つとして加齢とともに体内に蓄積した老化細胞が関与している可能性が最近指摘されている。老化細胞とは生体内で「細胞老化」をおこして不可逆的に細胞増殖を停止した細胞であり、様々な炎症性蛋白質を分泌することが知られているが、我々は老化細胞が炎症性蛋白質だけではなくエクソソームの分泌も亢進していることを見出し、加齢性疾患とエクソソームの解析を続けている。
本稿では、老化細胞が分泌するエクソソームに関する我々の研究を中心に最新の知見について概説したい。
細胞老化とは
一部の組織幹細胞を除いて、我々の体を構成している細胞の多くは細胞分裂の回数に限界があり、正常な体細胞が分裂寿命を迎えて不可逆的に増殖を停止した状態は細胞老化とよばれている 1)。また、若い正常な細胞に発がんの危険性のあるストレス(テロメアの短小化、がん遺伝子の活性化、酸化的ストレスなど)が加わった場合にも、やはり細胞老化が誘導され細胞の増殖は不可逆的に停止することから、細胞老化はアポトーシスとならんで異常が生じた細胞が増殖するのを防ぐ重要ながん抑制機構として働いていると考えられている 2)。
しかし、老化した細胞は細胞死とは異なり生体内で長期間生存し続けることから、加齢とともに体内に老化細胞が蓄積してゆくことも明らかとなっている 3)。一方で、老化細胞は持続的な DNA 損傷応答によってクロマチン構造が変化しており、炎症性サイトカイン・ケモカイン・マトリックス分解酵素や増殖因子のような様々な炎症性蛋白質の遺伝子発現が活性化している 4)。
そして、老化細胞はそれらの炎症性蛋白質を細胞外へと分泌することが知られており、このような細胞老化の表現型は SASP(Senescence-associated secretory phenotype)と呼ばれている。老化細胞から分泌された SASP 因子は周囲の組織に慢性的な炎症を引き起こすことから、あらゆる加齢性疾患の要因となっている可能性が示唆されている 5)。
我々は老化細胞が分泌する SASP 因子の解析を行う過程で、老化細胞では細胞外分泌膜小胞の一種であるエクソソームの分泌も亢進していることを見出した 6)。
エクソソームとは
エクソソームは、後期エンドソームである多胞性エンドソーム(Multi vesicular body)を経由して細胞から分泌される直径 50-150nm 程の細胞外膜小胞であり、蛋白質、脂質、核酸のような様々な細胞内構成分子がエクソソームに含まれて他の細胞に伝達することから、細胞間コミュニケーションに働いていることが近年報告され大変に注目を集めている 7)。
細胞老化とエクソソーム
ヒトの正常線維芽細胞を用いて分裂寿命による細胞老化(Replicative senescence)と活性化型がん遺伝子(RasV12)による細胞老化(Oncogeneinduced senescence)を誘導し、細胞が分泌するエクソソームの量を NTA法(Nanoparticle Tracking Analysis)で計測したところ、老化細胞ではエクソソームの分泌量が若い細胞の約 30 倍亢進していることが明らかとなった 6)。
そこで、我々は老化細胞で分泌が亢進しているエクソソームの生物学的機能に着目し解析を行った。老化細胞でエクソソームの生合成と分泌に重要な Alix と Rab27a をそれぞれノックダウンしエクソソーム経路を阻害したところ、アポトーシス様の細胞死が誘導された。驚くべきことに、老化細胞だけではなく若い増殖中の正常細胞においても、エクソソーム経路の阻害は DNA 損傷応答の活性化と細胞死を誘導したことから、正常な細胞はエクソソーム経路によって何か有害な物質を細胞外へと分泌している可能性が示唆された。
そこで、エクソソームに含まれて分泌され正常細胞に DNA 損傷を誘発する分子を探索したところ、それが細胞質のゲノム DNA 断片であることを見出した。細胞質に存在する DNA 断片は細胞質 DNA センサー(cGAS-STING 経路)によって感知され、自然免疫応答を引き起こすことが知られている。つまり、正常な細胞はエクソソームによって細胞質の有害なゲノム DNA 断片を細胞外へと分泌し、自己 DNA に対する自然免疫応答を防いでいると考えられる。
さらに、エクソソームはアデノウイルスなどの外来性の DNA に対してもウイルス感染のバリアとして働いていることから、エクソソーム経路は細胞の恒常性を維持するための重要な生体防御機構の一つであることが示された 6)。
また正常な細胞では、細胞質に存在する DNA 断片は DNase2a や TREX1 のような DNA 分解酵素によって速やかに除去されることが知られているが、我々は老化細胞では DNA 分解酵素の発現レベルが低下することを見出した 8)。そのため、老化細胞では細胞質に蓄積したゲノム DNA 断片が DNAセンサー経路の活性化を介して自然免疫応答を引き起こし、SASP を誘導することで肥満誘導性の肝がんの発症に関与していることを報告した(図1)。
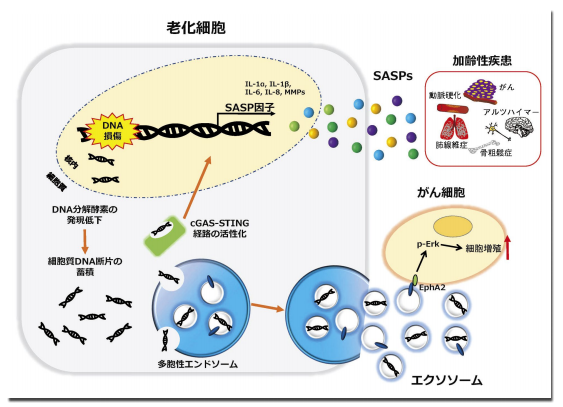
図1.老化細胞とエクソソーム
SASP としてのエクソソーム
がん微小環境中の老化細胞は SASP因子を介してがん細胞の増殖や転移・浸潤を助長する CAFs(Cancer-associated fibroblasts)として機能することが知られている。我々は老化細胞が分泌するエクソソームの機能を調べるために、若い細胞と老化した細胞から回収したエクソソーム画分をそれぞれ乳がん細胞株(MCF-7)に加えたところ、老化細胞が分泌したエクソソーム画分のみでがん細胞の増殖促進作用が観察された。
次に、それぞれの細胞から回収したエクソソーム画分のプロテオーム解析を行った結果、老化細胞が分泌したエクソソームではチロシンキナーゼ型受容体の一つでありがん転移促進因子として知られる Ephlin-A2(EphA2)の含有量が顕著に増加していることが明らかとなった。そして、老化細胞では EphA2 の脱リン酸化を制御する PTP1B が失活しているためにリン酸化された EphA2 が選択的にエクソソームへ取り込まれるというエクソソームの積荷の選択性を決定するメカニズムがあることを見出した。
そのため、老化細胞が分泌したエクソソーム膜上の EphA2 が、がん細胞で高発現している受容体(Ephlin-A1)と結合し、下流の Erk シグナルが活性化することで、がん細胞の増殖が促進することが示された。この研究の中で我々は、老化細胞由来の細胞外分泌膜小胞が、がん細胞の増殖促進を促すSASP 因子として働くことを明らかにした 9)。
おわりに
エクソソームはこれまで、その細胞間コミュニケーションツールとしての機能や診断ツールとしての有用性が特に注目され研究が進んできたが、細胞が何故エクソソームを分泌するのか、その生物学的な意義についてはまだまだ未知の部分が多い。
最近、視床下部幹細胞と前駆細胞を除去した早老症モデルマウスを用いた解析によって、視床下部幹細胞に由来するエクソソーム中の miRNA 量を調節することで加齢性疾患の発症速度と個体の寿命が変化することが報告されている 10)。さらに治療効果のある特定の siRNA やmRNA をエクソソームに積み込み標的細胞へと送り込む技術が次々と開発されていることから 11, 12)、今後エクソソームは単に疾患のバイオマーカーになるだけでなく、有用な DDS(Drug delivery system)として利用できる可能性が期待される。
参考文献
- Hayflick, L. : Exp. Cell Res., 37, 614 (1965). DOI: 10.1016/0014-4827(65)90211-9
- Takahashi, A. et al. : Nat. Cell Biol., 8, 1291 (2006). DOI: 10.1038/ncb1491
- Yamakoshi, K. et al. : J. Cell Biol., 186, 393 (2009). DOI: 10.1083/jcb.200904105
- Takahashi, A. et al. : Mol. Cell, 45, 123 (2012). DOI: 10.1016/j.molcel.2011.10.018
- He, S. et al. : Cell, 169, 1000 (2017). DOI: 10.1016/j.cell.2017.05.015
- Takahashi, A. et al. : Nat. Commun., 8, 15287 (2017). DOI: 10.1038/ncomms15287
- Tkach, M. and Théry, C. : Cell, 164, 1226 (2016). DOI: 10.1016/j.cell.2016.01.043
- Takahashi, A. et al. : Nat. Commun., 9, 1249 (2018). DOI: 10.1038/s41467-018-03555-8
- Takasugi, M. et al. : Nat. Commun., 8, 15729 (2017). DOI: 10.1038/ncomms15728
- Zhang, Y. et al. : Nature, 548, 52 (2017). DOI: 10.1038/nature23282
- Kamerkar, S. et al. : Nature, 546, 498 (2017). DOI: 10.1038/nature22341
- Kojima, R. et al. : Nat. Commun., 9, 1305 (2018). DOI: 10.1038/s41467-018-03733-8
関連記事




関連コンテンツ




